心情随笔
参与小麦穗发育的lncRNA 的全基因组分析


lncRNA:
长非编码RNA(long non-coding RNA,lncRNA)是一类RNA,其长度大于200个核苷酸,无能编码蛋白质。尽管lncRNA不能编码蛋白质,但它们在基因表达调控和其他生物学过程中发挥了重要作用。lncRNA可以在基因转录调控、表观遗传学调控、信号途径、细胞周期调节等方面发挥重要作用。lncRNA的生物功能包括:染色质重塑、基因转录调控、RNA的代谢和转运、蛋白质稳定性的调控、细胞周期调控、信号转导通路的调控等。与蛋白质编码RNA(protein-coding RNA)相比,lncRNA具有低保守性、高表达动态范围和组织特异性表达等特点。这些特征使得lncRNA在疾病的发生和发展中也扮演了重要的角色,例如癌症、心血管疾病和神经系统疾病等。总之,lncRNA作为一类新型的RNA,其生物功能的研究对于理解生命活动的调控机制以及疾病的发生和发展具有重要的意义。
文章主要内容:
在这篇文章中,作者分析了发育中的小麦穗的动态转录组,鉴定出了共计8,889个表达的lncRNA,其中在不同发育阶段有2,753个差异表达(DE)的lncRNA。作者进一步预测了315个DE lncRNA-mRNA调控对,并分析了这些DE lncRNA所调控的分子事件。随后,鉴定出了24个重要的lncRNA及其潜在的调控基因。最后作者还预测了一些lncRNA可能作为miRNA的潜在前体或者是miRNA的靶向序列。
转录组比对、转录本组装和 lncRNA 鉴定:
作者收集了小麦在6个发育阶段(S1-S6)的穗,并对这些穗进行测序,每个发育阶段有三组重复样品。完成转录组测序后,作者首先去除了有污染和低质量的reads。然后,将每个样品的15 Gb干净的序列比对到Triticum aestivum L.参考基因组组装中,使用HISAT软件和默认参数。接下来,使用Stringtie软件组装转录本,并使用Gffcompare软件将这些转录本与已知的基因注释模型进行比较,以预测新的转录本。使用LGC和CNCI软件并设定默认参数,鉴定哪些长度大于200 bp的并且不具备蛋白编码功能的新转录本。最后,使用Pfam数据库确保预测的转录本没有蛋白质编码区域。通过三种判断方法的一致性,确定转录本是否为lncRNA。
LncRNAs 的表达分析:
随后使用FPKM方法计算基因在每个样品中的表达水平,其中具有平均FPKM≥1的lncRNAs和编码基因被认为是表达的。然后使用Circos软件在21条染色体上对鉴定到的lncRNAs进行可视化。DEseq2用于在鉴定在不同的发育阶段有表达差异的lncRNAs和编码基因,FPKM表达量最少有两倍表达差异且p值小于0.05的DE lncRNAs和DEGs会被认为是差异表达的基因。随后使用k-means函数对DE lncRNAs进行聚类分析。
LncRNA 靶基因预测及 GO 分析:
预测长链非编码RNA(lncRNA)和其调控的编码基因之间的相互作用关系,如果lncRNA与编码基因重叠或距离不超过100 kb,则认为这些编码基因可能是lncRNA的顺式调控基因。使用LncTar计算lncRNA-mRNA对的自由能,并设置归一化自由能为-0.1的阈值来确定靶基因。通过Pearson相关系数(|r|> 0.9)过滤lncRNA-mRNA对,并将被DE lncRNAs调控的差异表达基因(DEGs)作为进一步分析的候选编码基因。使用TBtools软件进行GO富集分析,p值低于0.05的GO项被认为具有显著富集。
预测miRNA的前体和潜在靶标:
使用BLAST工具将小麦长链非编码RNA(lncRNA)序列与miRBase中的miRNA前体序列比对,筛选出相似性为100%且e-value<0.001的潜在miRNA前体。然后,使用psRNAtarget工具将小麦成熟miRNA序列与lncRNA序列比对,预测出lncRNA潜在靶向的miRNA.
Fig. 1
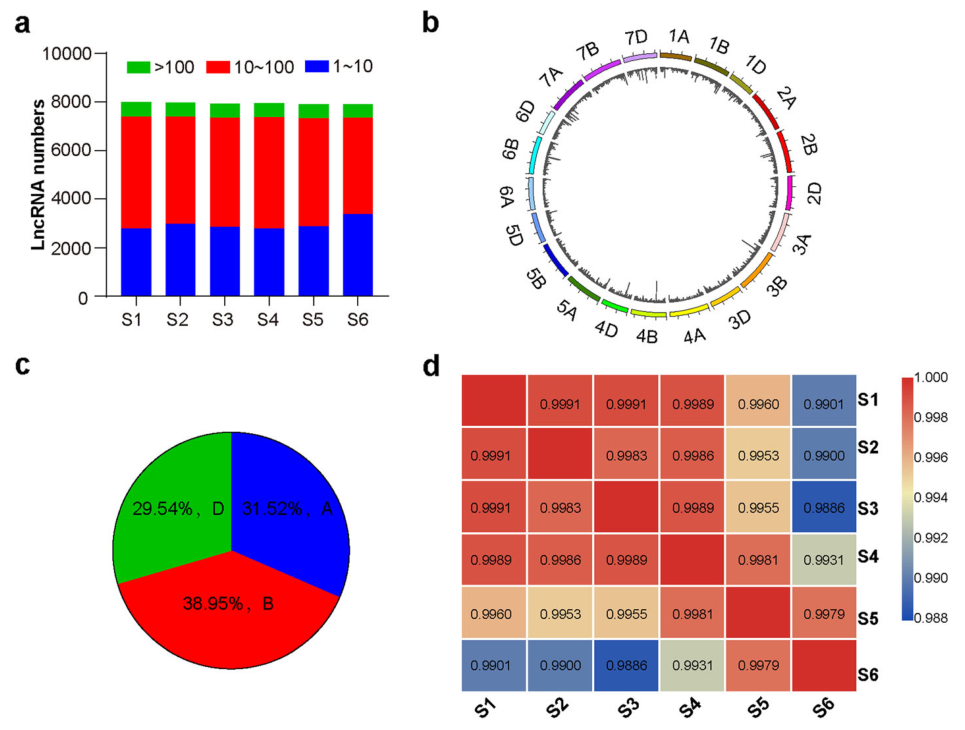
a 图展示了在小麦发育过程中不同阶段检测到的表达的 lncRNAs 数量,其中 S1-S6 分别代表不同的的发育阶段。不同表达量(TPM)的 lncRNAs 以不同颜色显示。b 图显示了表达的 lncRNAs 在小麦染色体上的分布情况。c 图展示了 lncRNAs 在小麦 A、B 和 D 三种同源染色体上的分布情况。d 图展示了从 S1 到 S6 六个发育阶段检测到的 lncRNAs 之间的相关系数。
Fig. 2
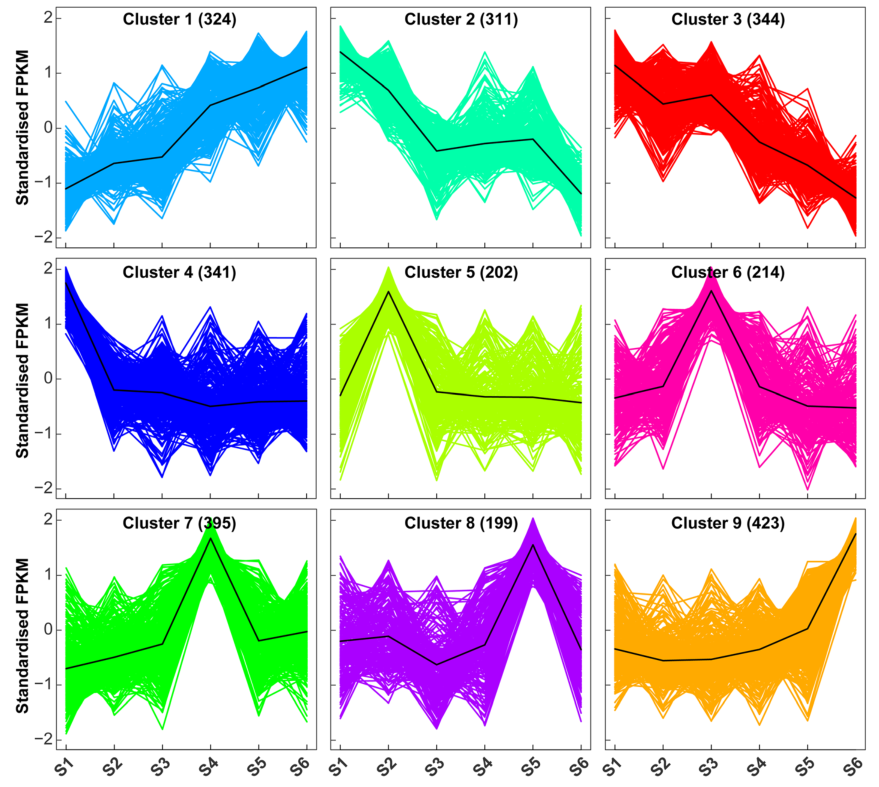
不同表达的长链非编码RNA聚类分析。聚类1代表从S1到S6一直表达升高的长链非编码RNA。聚类2和3包含从S1到S6一直表达下降的长链非编码RNA,具有不同的模式。聚类4-9分别代表在S1-S6中特定发育阶段丰富表达的长链非编码RNA。括号中的数字表示相应聚类中的长链非编码RNA数量。所有差异表达的长链非编码RNA(2,753个)的标准化FPKM值按欧几里得距离进行k均值聚类(k = 9)
Fig. 3
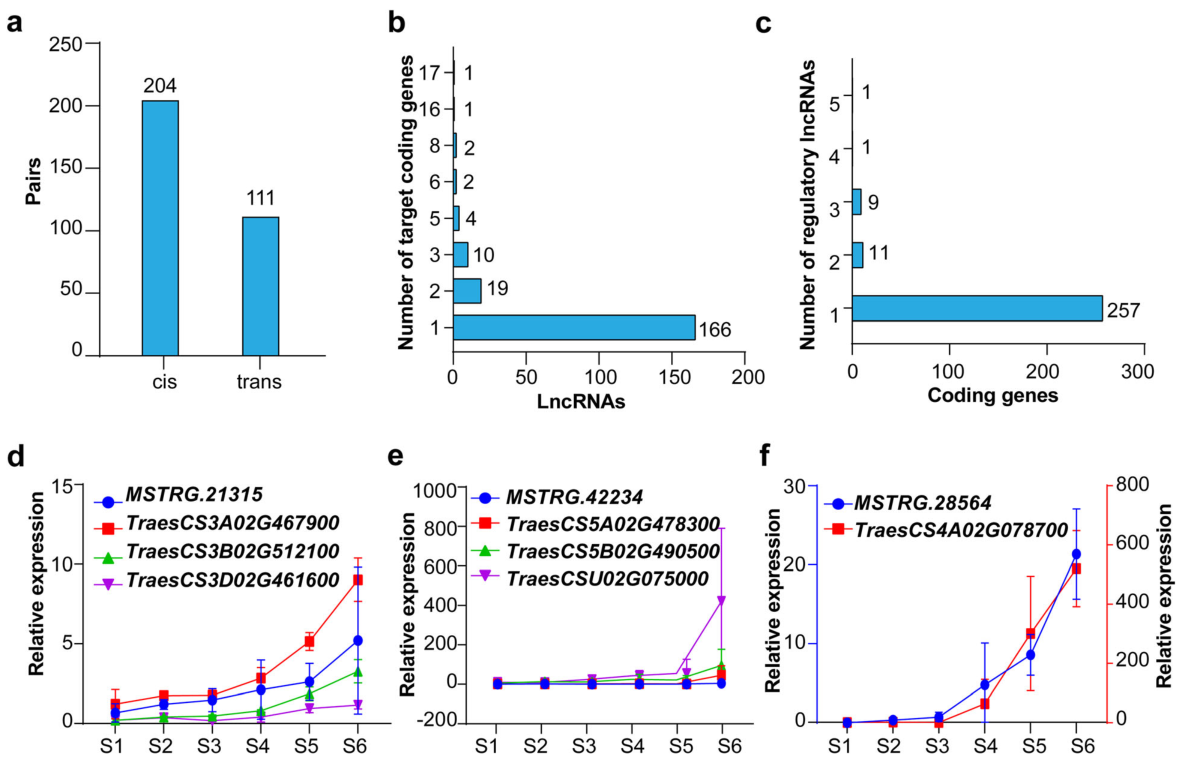
Fig. 4
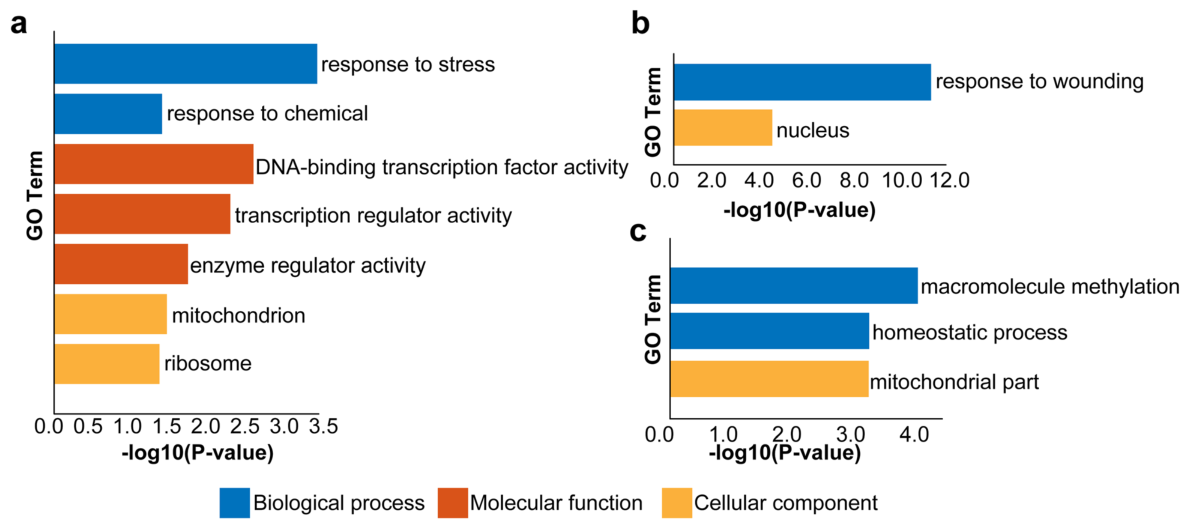
被差异表达的lncRNA靶向的差异表达基因的GO富集分析结果。a 被差异表达的 lncRNA 以cis-或trans-调控的差异表达基因的GO富集分析结果。 b 被表达量持续增加(从S1到S6)的 lncRNA 以cis-或trans-调控的差异表达基因的GO富集分析结果。c 被表达量持续降低(从S1到S6)的 lncRNA 以cis-或trans-调控的差异表达基因的GO富集分析结果。GO富集分析结果分为三个主要类别:生物过程、分子功能和细胞组成。P值<0.5的GO术语被鉴定为显著。
Fig. 5

由于被阶段特异性lncRNA靶向的编码基因可能有助于在个别发育阶段内的穗形成,因此我们还研究了聚类4-9中被lncRNA靶向的基因(图5;补充表S8)。正如预期的那样,这些编码基因确实在相应的发育阶段表现出优势表达,就像它们的配对lncRNA一样(图5)。在S1,有四个编码代谢相关蛋白质的基因,三个编码功能未知的Ycf68蛋白质,多个编码细胞色素、谷氨酸受体、含有五肽重复的蛋白质和锌指蛋白质的基因(图5a)。在S2,三个lncRNA参与调控编码一个触发因子、一个核糖体蛋白和一个生长素响应因子的三个基因(图5b)。在S3,编码胰蛋白酶抑制剂和糖或脂代谢相关蛋白质的基因被鉴定为lncRNA的潜在靶点(图5c)。在S4期间,大部分编码伤口应答蛋白、脱水素蛋白和早期根瘤素相关蛋白的基因被发现受到lncRNA在靶向调控(图5d)。同样,在S5期间,33个被lncRNA靶向调控的基因中,有22个编码伤口应答蛋白(图5e;补充表S8)。这个观察结果表明,lncRNA调控的应激反应,特别是伤口反应,可能有助于S4和S5期间小穗、雄蕊和雌蕊原基的形成。在S6期间,被lncRNA靶向调控的基因中,有几个编码MADS-box转录因子、锌指蛋白和Argonaute1蛋白,还发现有大量的基因编码核蛋白,例如染色质相关蛋白(图5f),这意味着lncRNA调控的转录调控和表观遗传修饰对花器官发育至关重要。
Table 1
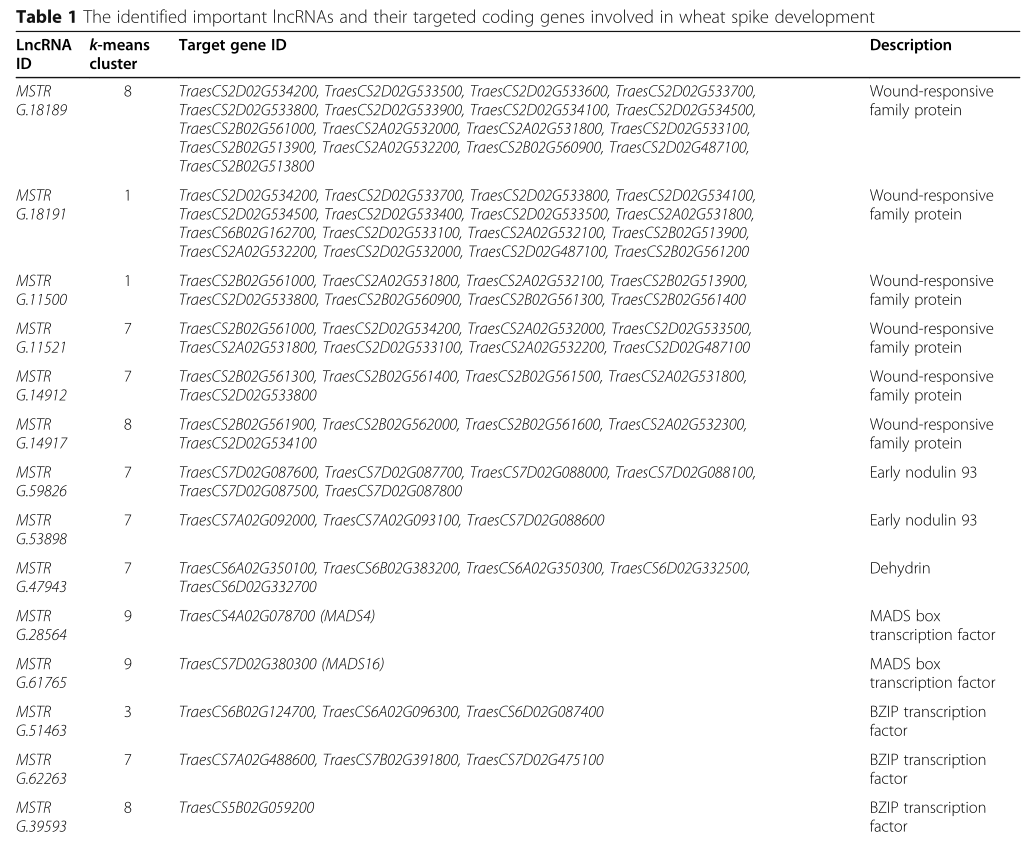
为了鉴定参与穗发育的重要 lncRNAs,作者仔细研究了与应激反应、TF 调节、激素信号传导和染色质重塑相关的 DE lncRNAs (表1)
Fig6.
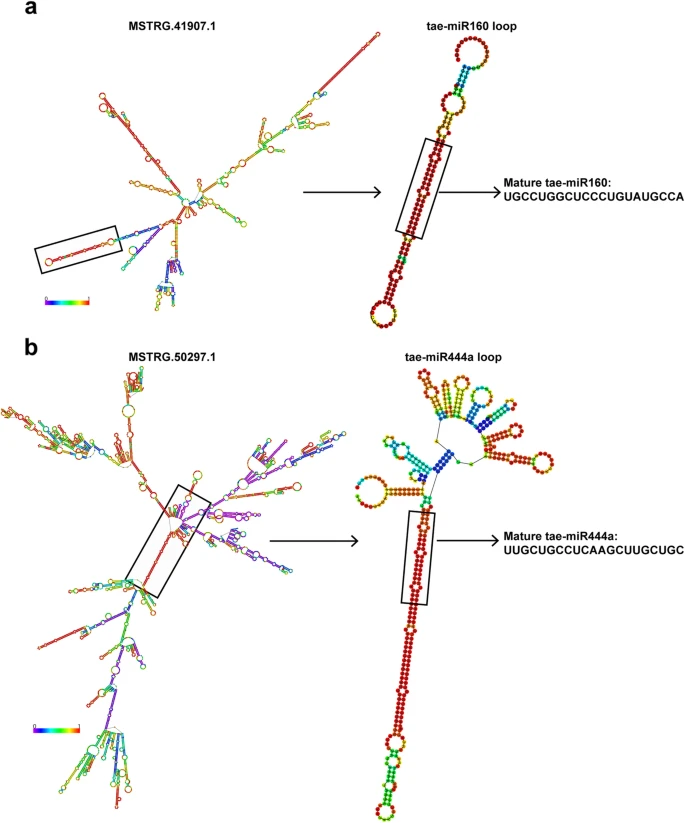
LncRNA 也可以作为 miRNA 的前体或靶标模拟物来调节 miRNA 的功能,据报道一些 miRNA 参与小麦穗发育,如 miR172和 miR156。为了鉴定可能作为 miRNA 前体或靶标模拟物的 lncRNA,作者进一步对 miRBase 和 psRNA 靶标进行了序列比对。共有58个 DE lncRNA 被预测为21个 miRNA 的潜在的前体。为了进一步验证这一点,作者使用 RNAfold Web 服务器,以预测两个具代表性的 lncRNAs 及其 miRNA 前体的二级结构。如图6a 所示,MSTRG.41907.1的二级结构表现出多个茎环结构,其中一个可以被切割以释放 tae-miR160的前体序列并最终形成成熟的 tae-miR160。类似地,MSTRG.50297.1的二级结构可能被切割以产生 taemiR444a 的前体并形成成熟的 tae-miR444a (图6b)。


最新文章
推荐阅读
网站浏览
